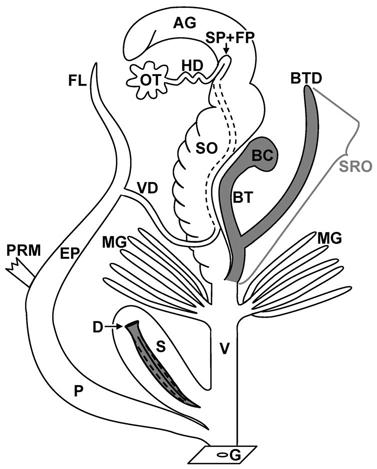
Schematic illustration of the genital apparatus: Brown garden snail (Cornu as-
persum).
Source: Koene, Schulenburg: BMC Evolutionary Biology 2005, 5:25.
|
Schematic illustration of the genital apparatus: Brown garden snail (Cornu as- persum). Source: Koene, Schulenburg: BMC Evolutionary Biology 2005, 5:25. |
The organ system in a Roman snail's body, which is called the genital apparatus, does not exclusively contain genital organs, but also different auxiliary organs, that have various tasks to perform depending on the stage of mating and reproduction. The genital opening is located behind the head on the right side of the body. From here the genital apparatus' organs reach until deep in the visceral sac. In the very rare left handed mutants also called snail kings, the genital apparatus opens on the left side of the body.
On the first sight, the genital apparatus appears to be built very complicatedly. That is why here the single organs shall be described according to their function during different phases of mating.
Hermaphroditic parts:
AG: Albumen gland; OT: Ovotestis or hermaphroditic gland; HD: Hermaphroditic
duct; SP + FP: Seminal pouch and fertilisation pouch; SO: Spermoviduct; G:
Atrium.
Female parts:
BC: Bursa copulatrix; BTD: Bursa tract diverticulum; MG: Finger shaped mucus
glands; V: Vagina; S: Dart sac; D: Love dart. SRO means
spermatophore receiving organ.
Male parts:
VD: Vas deferens (Spermatic duct); FL: Flagellum; EP: Epiphallus; P: Penis; PRM:
Penis retractor muscle.
A Roman snail's germ cells, sperm cells and egg cells, originate in the same hermaphroditic gonad, the hermaphroditic gland or ovotestis. Egg cells and sperm cells basically develop the same way. Wall cells in the gonad differentiate into mother cells from which develop, after several mitoses and on meiosis, different intermediate stages and finally the germ cells. As usual in nature, there are very many sperm cells, but only very few egg cells.
In contrary to the egg cells, that leave the ovotestis only shortly before fertilisation, sperm cells are produced continuously and released into the hermaphroditic duct. There they are stored in an immobilised state in wall pouches called seminal vesicles. When they are needed, the sperm cells migrate under their own power from the hermaphroditic duct into the spermoviduct.
In the spermoviduct the spermatic duct is formed like an open groove attached to the oviduct. To prevent auto-fertilisation, the connection is closed during copulation, so the sperm cells can only follow the spermatic duct. There is also a gland attached to the spermatic duct that is called prostate or spermatic duct gland. It produces a secretion that provides the passing sperm cells with energy and nutrients. It also immobilises them, so from here the sperm cells are transported by peristaltic motions of the spermoviduct.
Further to the exit the spermoviduct splits into the oviduct, which opens into the vagina, and on the other side into the spermatic duct as such, also called the vas deferens (literally the efferent duct), which opens into the so called epiphallus, where the sperm cells are transported further.
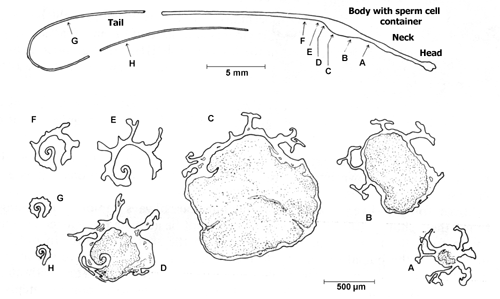 The Spermatophore. The cross-sections D to G show very well, that the spermatophore is not closed, so the sperm cells can escape after transfer. Source: Lind (1973). |
As mentioned already, transfer of sperm cells during copulation of one snail with another happens by the aid of a so-called spermatophore, a specialised means of transport. While the sperm cells are approaching, the spermatophore is produced in the area, where the vas deferens leads into the epiphallus. Gland cells in the epiphallus secrete matter to build the spermatophore that solidifies on the epiphallus' walls and so receives their surface structure.
The structure remains open where the vas deferens opens into the epiphallus, whereas it is closed on all sides further to the front. The spermatophore's thread-like tail is formed in the flagellum. This whip-shaped organ starts at the epiphallus' end. A longitudinal fold causes the spermatophore tail being open at the bottom.
As soon a the mass of prostate fluid containing sperm cells leaves the vas deferens opening, the spermatophore begins moving forward. So the opening in the spermatophore body is closed behind the sperm cells. Even further in front, the spermatophore's frontal part is formed into a head-neck-structure by peristaltic motions of the penis wall.
In the picture above, sections G and H are supposed to be 4 and 8 cm away from the spermatophore body proper, so the spermatophore of Helix pomatia is about 11 cm long including its tail.
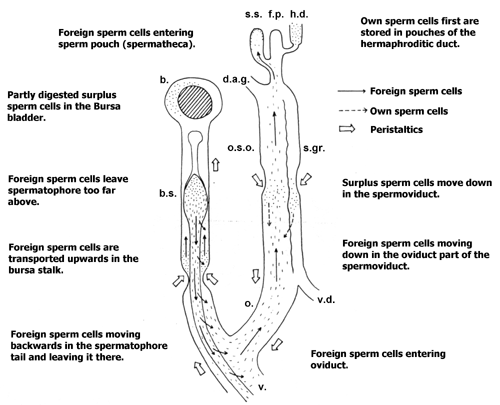 Illustration: The sperm cells' path after copulation. Legend: b.: Bursa copulatrix pouch; b.s.: Bursa stalk; d.a.g: Albumen gland duct; s.s.: Sperm pouch; f.p.: Fertilisation pouch; h.d.: Hermaphroditic duct; o.s.o.: Oviduct part of the spermoviduct; s.gr.: Seminal groove (Spermatic duct); o: Oviduct as such; v.d.: Vas deferens. Source: Lind (1973). |
The spermatophore with the sperm cells in it now is transferred using the penis into the mating partner's genital apparatus. There it is placed in a side channel of the vagina, which has a bladder-shaped pouch at its end, the bursa copulatrix (Copulation pouch).
For a long time, the function of the bursa copulatrix was unknown. In principle it was assumed the bursa copulatrix served to collect sperm cells until fertilisation. That is why it has been called receptaculum seminis (literally: "Sperm receiver") in many older pictures.
Actually the role of the bursa copulatrix is quite the contrary. It does receive the sperm cells, but to digest them by enzymes. Against that background, the spermatophore not only is used to transport the sperm cells, but also to protect them. So when the head and neck of the spermatophore continuously are moved towards the bursa by peristaltic motions, the sperm cells can leave the spermatophore through the backward opening. This also answers the question, of how nature could develop a transport medium which is open on the bottom.
The peristaltic motions of the bursa stalk would however not only transport the spermatophore, but also a great deal of sperm cells to certain death in the bursa's enzymatic fluids. With nearly 10 cm length, the spermatophore tail however is considerably longer than the bursa stalk, so some sperm cells leaving the spermatophore are almost at the entrance to the oviduct and out of danger.
In spite of the spermatophore supporting them, usually only 1 of sperm cells manages to escape the bursa copulatrix into the oviduct.
So what is the biological sense of an organ like the bursa copulatrix that renders several hours of courtship and mating useless by destroying the sperm cells just transferred? To understand, one has to take into consideration, that in sexual reproduction, there always are very few eggs cells against very many sperm cells. In front of this background it becomes obvious that it must be in the interest of the mating partner who just received a load of sperm cells that only the fittest of those are allowed to fertilise any egg cells. That explains the need for a means of selection, which is where the bursa copulatrix comes in. Only the few fittest sperm cells that manage to escape the draw of the bursa and to enter the oviduct are deemed fit for fertilisation.
In addition to foreign sperm cells and spermatophores the bursa copulatrix also recycles unused own sperm cells, as well as prostate fluid. This is why the connection between oviduct and seminal groove remains open, except during the actual copulation, when all the own sperm cells are supposed to reach the spermatophore.
Not only the acceptor, but also the donator of sperm cells, has the possibility to change conditions a bit to his favour. This is where the love dart comes in. Actually, rather than serving as a mere tool of excitation, the love dart actually is a means of injection, that transfers a secretion containing hormones that is produced by the finger shaped glands in the donator's genital apparatus. Those hormones cause a dilatation of the bursa stalk, so more sperm cells can escape. Further, they cause peristaltic motions of the oviduct, which support the sperm cells migration into the spermoviduct.
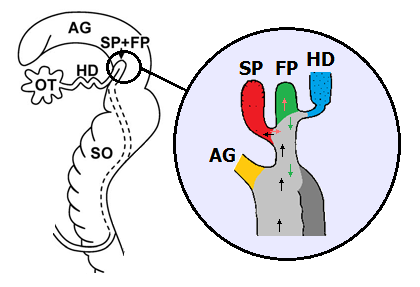 Seminal pouch (SP) and fertilisation pouch (FP) in Cornu aspersum. Left: Situation in the genital apparatus. Enlarged image: Yellow: Albumen gland duct. Red: Seminal pouch. Green: Fertilisation pouch. Blue: Hermaphroditic duct. Path of the sperm cells: Black: On the way to the seminal pouch. Light red: From the seminal pouch to the fertilisation pouch. Path of the fertilised egg cells: Green. Light grey: Spermoviduct. Dark grey: Spermio- duct (only separated partly). Further abbreviations: See above. Sources: Koene, Schulenburg: BMC Evolutionary Biology 2005, 5:25 (left) and Lind (1973) (right). Rearranged by R. Nordsieck. |
The few surviving sperm cells now have a long way to go towards the end of the spermoviduct. Here, lying in the large albumen gland of the snail, there is the fertilisation pouch. An especially detached blind side pouch serves as seminal pouch or spermatheca. Here all sperm cells from all copulations that a snail managed to arrange are stored. For fertilisation, they only have to leave the seminal pouch and enter the neighbouring fertilisation pouch.
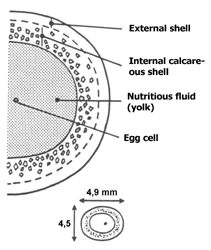 Cross-section through an egg of Cornu as- persum. Source: Tompa (1984). |
Only directly connected with the deposition of eggs, egg cells are transported from the ovotestis into the fertilisation pouch. There fertilisation takes place by sperm cells from the seminal pouch. When they leave the fertilisation pouch, the now fertilised egg cells (or zygotes) the large albumen gland provides them with a nutritious albumen coat. This nutritious coat, in spite of the gland's name, not only consists of albumen (egg white), but also of glyco-lipids and other nutrients.
The by now shell-less eggs from here are transported into the oviduct part of the spermoviduct, where they are put in line like pearls on a necklace. This part of the spermoviduct also is called the uterus, because here there are gland cells that provide the eggs with several coating layers, that may be quite rich in lime, and so also can be called an egg shell. X-Raying a snail during oviposition, one can see that the eggs become gradually less translucent towards the exit because of the growing content of lime in their shell.
Additionally to the lime shell, the Roman snail's eggs also receive a mucus coat that should protect them against fungal and other infections.
When they are deposited, the 40 to 60 eggs of a Roman snail have reached 7 to 8 mm in diameter.